Archives
- 2026-05
- 2026-04
- 2026-03
- 2026-02
- 2026-01
- 2025-12
- 2025-11
- 2025-10
- 2025-09
- 2025-04
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-07
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2019-07
- 2019-06
- 2018-07
-
g 15 br Material and methods br Results
2019-08-12
Material and methods Results Discussion In vitro evidence indicates that LAS191859 is a potent CRTh2 antagonist as demonstrated in GTPγS binding studies with potency in the low nanomolar range and in addition it demonstrates a long receptor residence time. This potency is also maintained in
-
Conclusions br Acknowledgements The study was supported
2019-08-12
Conclusions Acknowledgements The study was supported by a grant from the Argentine National Agency for the Promotion of Science and Technology (ANPCyT) (PICT 2012-2649). N.R.S., H.H.O., G.J.H. and F.R. are research career members and N.C.G., E.A., E. H., F.M.R., are fellows of the National Scien
-
Prostaglandin E receptor subtype EP is a transmembrane G cou
2019-08-12
Prostaglandin E receptor subtype 4 (EP4) is a transmembrane G-coupled protein receptor activated by prostaglandin E2 (PGE2). EP4 activation exerts anti-inflammatory effects in adipose tissue by dampening the levels of inflammatory chemokines [12]. In the mouse, EP4 deficiency aggravates fragmentatio
-
An increasing body of evidence demonstrates that MXC can
2019-08-09
An increasing body of evidence demonstrates that MXC can cause neurobehavioral and neuropathological alterations [7]. In the brain, there are several steroidogenic enzymes, which are responsible for neurosteroid production. Neurosteroids are subsets of steroids that rapidly change neuronal excitabil
-
Arsenic could cause apoptosis in both benign and
2019-08-09
Arsenic could cause apoptosis in both benign and malignant cells (Akao et al., 2000, Gupta et al., 2003, Namgung and Xia, 2001). However, at present, little is known about the consequences of arsenic-treated urothelial cells in autophagy and expression of its related proteins. In addition, the alter
-
Recently we have observed that
2019-08-09
Recently, we have observed that pharmacological PPARα and PPARβ/δ activation regulates the expression of genes involved in FA metabolism such as FAT/CD36, CPT1, LCAD and MCAD in Sertoli cells. We have also observed that PPARβ/δ activation can simultaneously regulate the expression of the above-menti
-
The following is the supplementary data
2019-08-09

The following is the supplementary data related to this article. Funding This work was supported by departmental funding to Prof. Mühl. Funding derived from the State of Hessia to the Institute of General Pharmacology and Toxicology at the pharmazentrum frankfurt (Head: Prof. Josef Pfeilschifter
-
cAMP mediated signaling pathways are important for maintaini
2019-08-09
cAMP-mediated signaling pathways are important for maintaining metabolic homeostasis, and the effects of the glucagon/catecholamine–cAMP–PKA axis on energy balance have been well documented [20]. For example, p m x of the PKA 2β regulatory subunit (RIIβ) in mice leads to increased expression of unc
-
Fatty acid oxidation plays a crucial role
2019-08-08
Fatty promotion info β-oxidation plays a crucial role in maintaining body energy homoeostasis mainly during catabolic states. It mainly occurs in mitochondria and involves a cyclical series of reactions that result in the shortening of fatty acids. Fatty acids comprise an efficient source of energy
-
Previous studies have documented that
2019-08-08
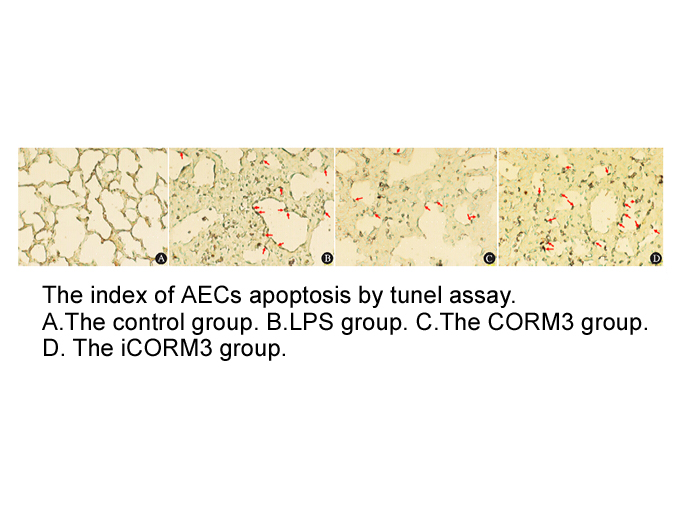
Previous studies have documented that SCFAs may elicit effects on lipid metabolism via de novo synthesis and transport. Acetate is not only used as an energy source, but is also constituted as a substrate for the cholesterol synthesis in the liver through acetyl-CoA, thus affecting plasma cholestero
-
br Acknowledgements We thank Dr Tai Yuan Yu and
2019-08-08
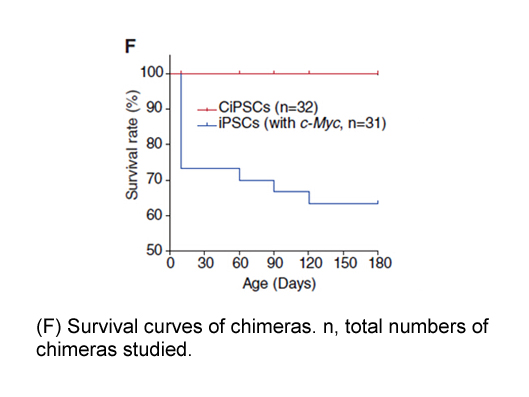
Acknowledgements We thank Dr. Tai-Yuan Yu and Miss Chun-Ping Chang for helping with the drug/DNA sequence specificity study. We also thank the Chemical Synthesis Core and the Pathology Core Laboratory of IBMS for synthesizing SL-1 and for performing the pathology analysis, respectively, and the T
-
Introduction The COMT gene is located on chromosomal
2019-08-08

Introduction The COMT gene is located on chromosomal band 22q11.2, extends over 28 kb, contains six exons, and encodes for catechol-O-methyl-transferase (Weinshilboum and Raymond, 1977; Grossman et al., 1992; Lundström et al., 1995; Bearden et al., 2005). This enzyme plays a major role in regulatio
-
Through the process of alternative splicing the
2019-08-08
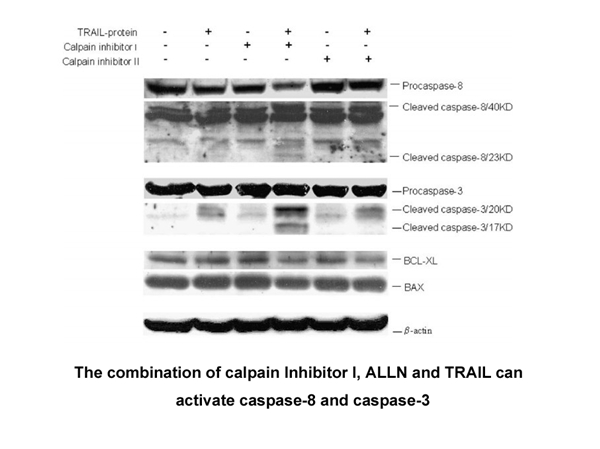
Through the process of alternative splicing, the IKZF1 gene is capable of encoding a large number of Ikaros isoforms (Molnar et al., 1996). Some of these isoforms were shown to have distinct functions (Li et al., 2011, Ronni et al., 2007). Ikaros protein contains four zinc fingers at the N-terminus
-
Notably HSP expression is under control of the tetrameric
2019-08-08
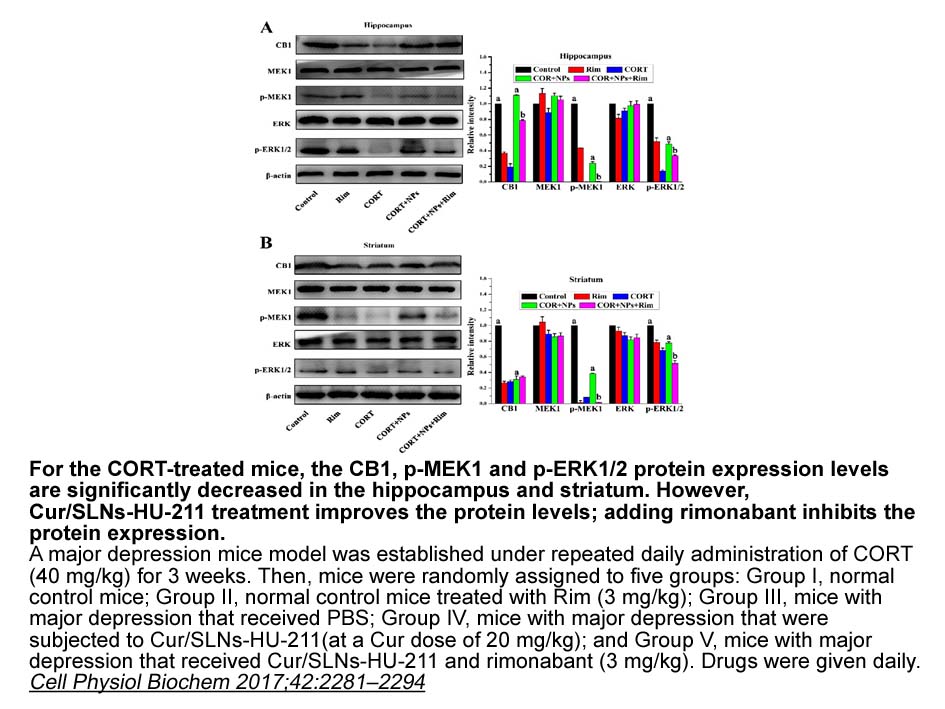
Notably, HSP27 expression is under control of the tetrameric form of CK2 holoenzyme, as knockout or knockdown of the β regulatory subunit alone reproduces the same effects of targeting the α/α′ catalytic subunits. This observation discloses the possibility to use more specific drugs that affects onl
-
In addition to PGC transcriptional co repressors
2019-08-08
In addition to PGC1α, transcriptional co-repressors such as NCOR and RIP140 participate in oxidative muscle remodeling induced by exercise, whereby reductions in their expression and the resulting de-repression of downstream TFs activates oxidative gene expression (Seth et al., 2007, Yamamoto et al.
15186 records 996/1013 page Previous Next First page 上5页 9969979989991000 下5页 Last page